Открыто новое кембрийское позвоночное
Находка кембрийского позвоночного — всегда событие.
Тогдашние позвоночные были мягкотелыми, в палеонтологической летописи они сохраняются редко. Между тем для понимания эволюции позвоночных крайне важны события кембрийского периода, когда их план строения находился в процессе становления. В данном случае находка позволила увидеть воочию один из промежуточных этапов процесса оформления головного конца тела позвоночных — цефализации.
Рис. 1. Nuucichthys: художественная реконструкция животного и его предполагаемое место на эволюционном древе. Иллюстрация из обсуждаемой статьи в Royal Society Open Science
Как выглядели первые позвоночные животные? Долгое время этот вопрос был чисто теоретическим. Достоверные остатки позвоночных были известны только начиная с ордовика, когда их план строения уже давно сложился. Было ясно, что какие-то ранние позвоночные должны были существовать и в кембрии, но про их облик палеонтология ничего не могла сказать.
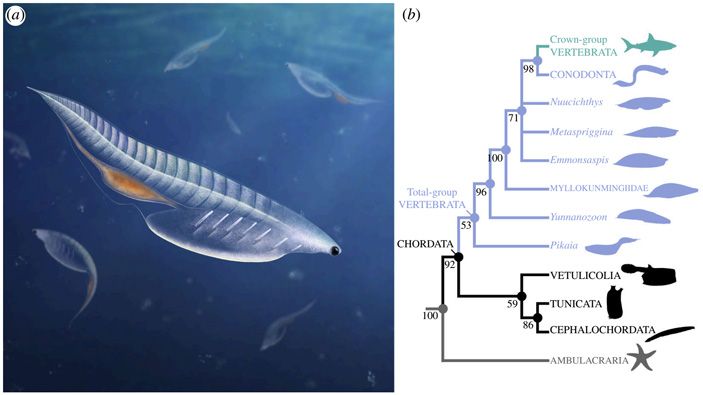
Оставалась сравнительная анатомия. Еще в первой половине прошлого века Алексей Николаевич Северцов, почти не используя палеонтологических данных, реконструировал общего предка всех хордовых животных, которого он назвал «примитивным бесчерепным» (Acrania primitiva). Его наиболее заметной чертой была сильная сегментированная мускулатура, причем сегменты мышц — миомеры — доходили до самого переднего конца тела. Это полностью соответствует тому, что мы видим у самого примитивного современного хордового — ланцетника. Такая мускулатура могла быть нужна только для плавания, поэтому общую форму тела «примитивного бесчерепного» Северцов реконструировал как рыбообразную. Центральная нервная система представляла собой расположенную на спинной стороне нервную трубку, под которой находилась упругая хорда. Передний отдел пищеварительного тракта — глотка — был пронизан парными жаберными щелями. У ланцетника число жаберных щелей очень велико (90 пар), но это явно вторичное увеличение; Северцов считал, что у «примитивного бесчерепного» число пар жаберных щелей было не больше 20. «Примитивные бесчерепные» были микрофагами, то есть питались мелкими органическими частицами, приносимыми с током воды. Вода шла через ротовое отверстие в полость глотки, там пищевые частицы отцеживались, а вода выходила через жаберные щели (рис. 2).
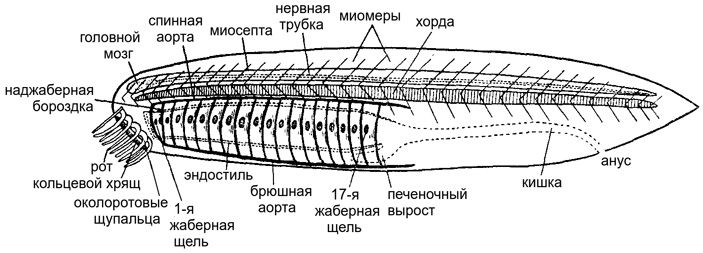
Рис. 2. Общий предок хордовых по Северцову. Число жаберных щелей, как указывает сам Северцов, дано лишь приблизительно. Эндостиль и наджаберная бороздка — части фильтрующего слизистого аппарата. Иллюстрация из книги А. Н. Северцов, 1967. Главные направления эволюционного процесса
Чтобы проверить эту модель, нужны были реальные находки кембрийских хордовых. А вот с ними в XX веке были большие трудности. Только в 1979 году британский палеонтолог Саймон Конвей Моррис (Simon Conway Morris) установил, что давно известная кембрийская пикайя (Pikaia), которую сначала приняли за кольчатого червя, на самом деле наверняка является хордовым животным: ее характерным образом исчерчивающие тело мышечные сегменты — это миомеры, которые не встречаются ни у кого, кроме хордовых. Какое-то время пикайя была единственным известным кембрийским хордовым. Увы, это не означало, что общий предок хордовых найден. Пикайя имеет целый ряд загадочных признаков, которые, скорее всего, свидетельствуют о ранней специализации в каком-то необычном направлении (J. Mallatt, N. Holland, 2013. Pikaia gracilens Walcott: stem chordate, or already specialized in the Cambrian?).
Загадки пикайи
Проблемы начинаются уже с ее внешнего вида (рис. 3, А). Голова пикайи несет впереди пару коротких щупалец, которые придают ей сходство с моллюском. В каждое щупальце входит нерв. Очевидно, функция щупалец была чувствительной. На что эти щупальца похожи? Их, конечно, можно сравнить с усами современных сомов или с чувствительными щупальцами миксин. Но и у тех, и у других морда всегда несет больше одной пары усов или щупалец, а у пикайи — только одну пару. С другой стороны, внешне довольно похожие чувствительные придатки есть у многих беспозвоночных: щупальца улиток, усики онихофор и членистоногих и тому подобное. Но при чем тут пикайя? Это остается загадкой.
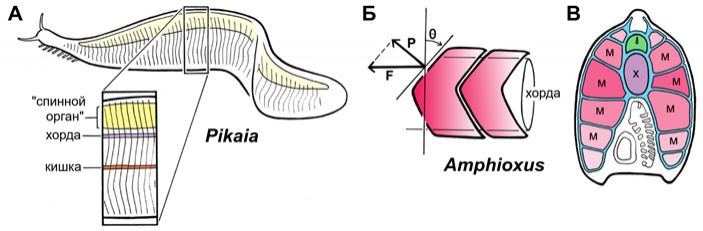
Рис. 3. Пикайя по сравнению с ланцетником. А — общий вид пикайи со щупальцами, миомерами и «спинным органом». Б — схема миомеров ланцетника, вид сбоку. Стрелки обозначают силы, которые создает миомер, сокращаясь. В — поперечный разрез туловища ланцетника. Обратим внимание на хорду (х) и миомеры (м). Благодаря тому, что миомеры перекрываются, на любом поперечном разрезе у ланцетника видна не одна пара миомеров, а сразу несколько. Соответственно и крепления миосепт распределены по хорде гораздо равномернее, чем если бы миомеры не перекрывались и миосепты были строго вертикальными. Иллюстрации из статьи T. Lacalli, 2012. The Middle Cambrian fossil Pikaia and the evolution of chordate swimming
Продолжая осматривать пикайю, мы увидим, что у нее очень маленький рот и совсем уж крохотные жаберные щели. Это тоже проблема. Дело в том, что все примитивные хордовые, не исключая и первых позвоночных, несомненно были фильтраторами: они прокачивали через глотку большое количество воды, выводили ее через жаберные щели, а мелкие органические частицы захватывали с помощью специального слизистого аппарата и отправляли дальше в кишечник. Так питаются оболочники, так питается ланцетник, так питается и личинка современной миноги — пескоройка; причем механизм фильтрационного питания у них у всех настолько сходен, что его общее происхождение не вызывает сомнений (J. Mallatt, 2023. Vertebrate origins are informed by larval lampreys (ammocoetes): a response to Miyashita et al., 2021). Но вот пикайя так питаться не могла. Ее крошечный рот и микроскопические жаберные щели просто не успевали бы пропустить нужный для этого объем воды. Она питалась каким-то другим способом. А поскольку никаких зубчиков или других твердых элементов ротового аппарата у пикайи нет, то, вероятно, она просто всасывала мелкие пищевые объекты, которые находила на дне, по одному за раз.
Вдоль брюшной стороны тела у пикайи проходит длинный тяж, который интерпретируется как брюшной кровеносный сосуд. Подвох тут в том, что подобный брюшной сосуд, идущий вдоль всего тела, часто встречается у беспозвоночных. А вот у позвоночных и у ланцетника главная вена, идущая вдоль брюха, всегда распадается на сетку капилляров — воротную систему печени. Потом эти капилляры сливаются в печеночную вену. Но у пикайи брюшной сосуд сплошной, ничего похожего на воротную систему печени не найдено, и что это за сосуд такой — неизвестно.
Строго говоря, единственный серьезный аргумент, заставляющий считать пикайю хордовым животным — это ее миомеры. Ленту осевых мышц, подразделенную на миомеры, трудно с чем-нибудь спутать, и ни у кого, кроме хордовых, мускулатура не бывает организована таким образом. Но и с миомерами у пикайи не все так просто. Сам по себе миомер — это блок мышечных волокон, направленных строго продольно, то есть спереди назад. Впереди и сзади мышечные волокна каждого миомера прикрепляются к миосептам, прочным перегородкам, которые, в свою очередь, прикреплены к оболочке хорды. Благодаря поочередному сокращению миомеров животное, собственно, и движется. Если теперь мы посмотрим на ланцетника, то увидим, что у него миомеры как бы входят друг в друга под острым углом, так что каждая миосепта образует V-образный контур. Зачем такие сложности и почему бы границам миомеров не быть прямыми?
Замечательный канадский биолог Тёрстон Лакалли (Thurston Lacalli) объясняет это следующим образом (T. Lacalli, 2012. The Middle Cambrian fossil Pikaia and the evolution of chordate swimming). Благодаря тому, что миомеры перекрываются, происходит выравнивание механических нагрузок на хорду и ее оболочку. Представим себе обратный случай: пусть границы миомеров будут строго вертикальными. Тогда в местах прикрепления миосепт к хорде возникнут очень узкие напряженные участки, за которые будут постоянно дергать мышцы, в то время как участки хорды между миосептами будут оставаться в покое. Поскольку миомеры сокращаются на правой и левой стороне не симметрично, хорда будет постоянно искривляться, ее оболочка будет морщиться, а в моменты особо сильных нагрузок она может и порваться. Если же миосепты будут изогнуты под углом (рис. 3, Б и В), то области их прикрепления к хорде будут гораздо более протяженными и нагрузки будут приходиться на хорду гораздо более равномерно, охватывая более-менее всю ее длину. И чем острее угол, тем сильнее будет этот эффект.
А теперь посмотрим на пикайю. У нее границы миомеров как раз вертикальны, с незначительными волнообразными изгибами (рис. 3, А). Это означает, что вся нагрузка на хорду будет приходиться на вертикальные крепления миосепт. Чтобы оболочка хорды в этих местах не порвалась, нагрузка должна быть резко ограничена. Отсюда — предположение Лакалли, что миомеры у пикайи были слабыми и скорость плаванья была низкой, совсем не такой, как у современного ланцетника или современной рыбы. Скорее всего, ее мышечные волокна были «медленными», физиологически неспособными быстро сокращаться: зачем, если скелет этого все равно не позволял?
Между тем во времена пикайи уже существовали крупные морские хищники, от которых она вряд ли могла быстро уплыть. Поэтому Лакалли считает, что она вела скрытный образ жизни и держалась почти все время около дна, ощупывая его своими усиками и подбирая мелкие пищевые частицы.
Но самая загадочная особенность пикайи — это «спинной орган», находившийся у нее прямо под покровом спинной стороны тела. Он довольно толстый («подобен сосиске», по выражению Лакалли), и его природа абсолютно непонятна. Этот орган никак не может быть хордой, потому что над хордой должна располагаться нервная трубка, а тут места для нее нет. «Спинной орган» представлял собой замкнутую трубку, заполненную какой-то внеклеточной массой. Возможно, это был гидростатический орган с полужидким содержимым, служивший или для упругости, или для плавучести, или для того и другого сразу. Но откуда он взялся? Тёрстон Лакалли предлагает несколько возможных сценариев образования этого органа (T. Lacalli, 2024. The Cambrian fossil Pikaia, and the origin of chordate somites), но сделать между ними выбор нельзя, потому что про эмбриологию пикайи мы не знаем ничего. Самая отчаянная версия заключается в том, что «спинной орган» — это просто-напросто нервная трубка, которая переполнилась жидкостью, раздулась и стала в придачу к нервным функциям служить механической опорой тела. Если не вдаваться в такие крайности, то можно вспомнить о спинном плавнике ланцетника, скелет которого — именно гидростатический: он образован серией плавниковых камер, маленьких замкнутых полостей, содержащих жидкость под давлением. Только вот у пикайи подобная камера одна, огромная, тянущаяся на все туловище. Никаких аналогов этого органа у современных животных мы, к сожалению, не знаем.
Забегая вперед, добавим, что пикайя — вовсе не самое древнее известное хордовое. Более того, она моложе некоторых позвоночных. Пикайя жила как минимум на 15 миллионов лет позже древнейшего известного «нормального» позвоночного — хайкоуихтиса (см. о нем ниже). Учитывая, что скорости эволюции в кембрии были очень высокими, это означает, что у пикайи хватало времени, чтобы специализироваться в какую угодно сторону. И от общего предка хордовых она, возможно, успела уйти очень далеко. В любом случае пикайя породила больше вопросов, чем дала ответов.
Первые позвоночные
Затем случилось то, что американский палеонтолог Марк Мак-Менамин (Marc McMenamin) назвал «китайским прорывом (Chinese Breakthrough). В середине 1980-х годов была открыта Чэнцзянская биота — кембрийская фауна уникальной сохранности, где удалось найти остатки множества небольших и бесскелетных животных (см. Палеонтологический заповедник Чэнцзян). Среди этих животных были и хордовые. Подобные открытия продолжались, и в результате сейчас известно примерно 11 родов кембрийских хордовых (M. McMenamin, 2019. Cambrian chordates and vetulicolians; оговорка «примерно» тут необходима, хотя бы потому, что некоторые из этих родов, возможно, являются синонимами друг друга). Но далеко не все они — позвоночные. Ведь, кроме позвоночных, к хордовым относятся бесчерепные (ланцетник и его древние родичи) и крайне своеобразные оболочники. Из одиннадцати родов кембрийских хордовых два рода относятся к оболочникам, три — скорее всего к бесчерепным, еще два — к юннанозоям, которых одни авторы считают позвоночными, другие бесчерепными, а третьи вообще отказываются относить их к хордовым. Вопрос о том, кто такие на самом деле юннанозои, невероятно запутан, и мы сейчас оставим его за кадром. Пикайя, как уже понятно, стоит особняком. И только оставшиеся три рода — это несомненные кембрийские позвоночные. Посмотрим, что же они собой представляют.
В 1999 году китайские палеонтологи описали сразу два маленьких рыбообразных существа, найденных в одном и том же кембрийском местонахождении (D. Shu et al., 1999. Lower Cambrian vertebrates from south China). Это были миллокунмингия (Myllokunmingia) и хайкоуихтис (Haikouichthys). Различия между ними касаются в основном формы тела и могут быть артефактами сохранности, поэтому некоторые палеонтологи считают, что миллокунмингия и хайкоуихтис — одно и то же (X. Hou et al., 2017. The cambrian fossils of Chengjiang, China: the flowering of early animal life). Хайкоуихтис известен по гораздо большему числу экземпляров, и большая часть полученной информации относится к нему (рис. 4).
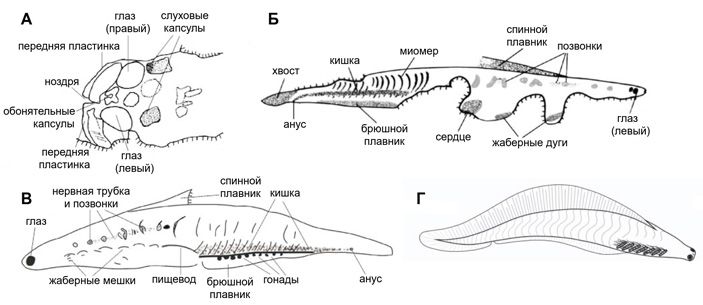
Рис. 4. Хайкоуихтис. А–В — рисунки хорошо сохранившихся ископаемых экземпляров, Г — реконструкция живого животного. Масштаб не соблюден. На реконструкции Г у хайкоуихтиса не 6, а 7 жаберных щелей — это допустимо, потому что их точное число так и остается спорным. А–В — иллюстрации из статьи D. Shu et al., 2003. Head and backbone of the Early Cambrian vertebrate Haikouichthys; Г — иллюстрация из статьи: X.-G. Zhang, X.-G. Hou, 2004. Evidence for a single median fin-fold and tail in the Lower Cambrian vertebrate, Haikouichthys ercaicunensis
Судя по хайкоуихтису, первые позвоночные были мелкими животными — длиной около 3 сантиметров. Вдоль тела хайкоуихтиса тянется ряд из примерно 25 типичных миомеров, причем контуры границ между миомерами сложные, в форме буквы W, а не простые в форме буквы V, как у ланцетника. Вместе с веретеновидной обтекаемой формой тела это показывает, что хайкоуихтис был достаточно хорошим пловцом. Никаких твердых покровных чешуй, щитков или пластин у хайкоуихтиса нет: это «голая рыба» («naked fish»), в отличие от панцирных позвоночных, которые появляются начиная с ордовика. Внутренний скелет у него исключительно хрящевой, если не считать хорды. Вдоль хорды регулярно расположены хрящевые структуры, которые могут быть только дугами позвонков, причем у хайкоуихтиса они развиты лучше, чем у современных бесчелюстных — у миноги и тем более у миксины. Таким образом, хайкоуихтис вполне заслужил право называться позвоночным животным.
Глотка хайкоуихтиса открывается наружу шестью парами жаберных щелей. На стенках жаберных щелей, судя по отпечаткам, сидели настоящие жабры — органы водного дыхания в виде расположенных рядами лепестков, пронизанных кровеносными капиллярами. Непосредственно позади глотки на отпечатках видны очертания околосердечной полости, и это значит, что у хайкоуихтиса было сердце. Эти признаки взаимосвязаны: сердце у позвоночных затем и появилось, чтобы прокачивать воду сквозь сеть жаберных капилляров, которые, как любые тонкие сосуды, обладают большим гидродинамическим сопротивлением.
На голове хайкоуихтиса хорошо заметны глаза. Судя по характеру отпечатков, они были камерными, со зрачком, хрусталиком и склерой. Между глазами находится пара небольших капсул, в которых скорее всего размещались органы обоняния. Позади есть отпечатки еще одной пары капсул — для внутреннего уха. Таким образом, у хайкоуихтиса был полный набор сложных органов чувств, характерных для позвоночных. Головной мозг не сохранился, но он, безусловно, был неплохо развит: чтобы обрабатывать информацию от таких органов чувств, нужны объемные нервные центры. Рядом находятся отпечатки хрящей, образующих мозговую коробку и ротовой аппарат, но о них нельзя в деталях сказать почти ничего, кроме того, что челюстей у хайкоуихтиса не было.
Интересная особенность хайкоуихтиса, оказавшаяся неожиданной для исследователей, состоит в том, что глаза у него находятся не по бокам головы, а на ее переднем конце. Такое же переднее расположение глаз наблюдается у самых древних панцирных бесчелюстных, которые появляются в ордовике. Раньше думали, что это какая-то специализация, но теперь похоже, что это чрезвычайно архаичная черта, которую ордовикские бесчелюстные унаследовали от своих кембрийских предков. В более поздние эпохи она исчезает.
С чем эта особенность может быть связана? По современным представлениям, истинный (морфологический) передний конец нервной трубки позвоночных находится где-то между перекрестом зрительных нервов и расположенной поблизости от него передней комиссурой (R. Nieuwenhuys, 2017. Principles of current vertebrate neuromorphology). С другой стороны, эмбриологи давно заметили, что, например, у зародыша аксолотля область формирования будущих глазных бокалов находится в самой передней части нервной пластинки, еще только превращающейся в нервную трубку (C. Jacobson, 1959. The localization of the presumptive cerebral regions in the neural plate of the axolotl larva). Эти данные указывают на одно и то же: некогда, у древнейших позвоночных, глаза находились рядом с передним концом мозга, а тем самым и всей головы. Их сдвиг назад произошел позже, в результате разрастания переднего мозга, связанного с органом обоняния. Как видим, палеонтологическая летопись подтверждает эти основанные на эмбриологических фактах догадки.
Орган обоняния у хайкоуихтиса тоже интересный. Обонятельные капсулы расположены между глазами, а не впереди от них, и очень близко друг к другу — настолько, что вход в них, судя по всему, был общим, то есть непарным (D. Shu et al., 2003. Head and backbone of the Early Cambrian vertebrate Haikouichthys). Непарная ноздря — признак, характерный для бесчелюстных позвоночных, к которым относятся миноги, миксины и многочисленные вымершие панцирные формы. В отличие от бесчелюстных, у рыб ноздри всегда парные. Палеонтологи долго спорили о том, какое из этих состояний более древнее (см. Л. И. Новицкая, В. Н. Каратаюте-Талимаа, 1986. Замечания о кладистическом анализе в связи с гипотезой миоптеригий и проблемой происхождения гнатостом), и хайкоуихтис, похоже, склоняет чашу весов в пользу бесчелюстных.
Теперь сравним хайкоуихтиса с общим предком хордовых, которого нарисовал около ста лет назад академик Северцов. Прежде всего хайкоуихтис относится к другому эволюционному уровню: «Acrania primitiva» — это бесчерепное, а хайкоуихтис — это позвоночное. У ланцетника, современного представителя бесчерепных, нет ни позвоночника, ни жабр, ни сердца, ни сложных парных органов чувств. Однако с учетом этой поправки можно сказать, что хайкоуихтис более-менее похож на реконструкцию Северцова. Примерный размер, форма тела, вероятный способ питания — все совпадает. На схеме Северцова изображено 34 миомера, у хайкоуихтиса их 25 — вполне сопоставимо. А вот число жаберных щелей Северцов преувеличил: судя по находкам, у предков позвоночных их никогда не было больше шести или семи пар. Но реконструкцию в целом это не разрушает.
В 2008 году уже упоминавшийся Саймон Конвей Моррис сделал описание загадочного кембрийского ископаемого под названием Metaspriggina, которое до этого исследовали лишь поверхностно (S. Conway Morris, 2008. A redescription of a rare chordate, Metaspriggina walcotti Simonetta and Insolm, from the Burgess Shale (Middle Cambrian), British Columbia, Canada). Метасприггина оказалась не только хордовым, но и позвоночным. В 2014 году Конвей Моррис и Жан-Бернар Карон (Jean-Bernard Caron) опубликовали подробное описание всех экземпляров метасприггины, какие только удалось найти (S. Conway Morris, J.-B. Caron, 2014. A primitive fish from the Cambrian of North America). Выяснилось, что метасприггина во многом похожа на хайкоуихтиса. Правда, она крупнее: длина наиболее сохранившихся экземпляров — около 6 сантиметров, а миомеров у них больше 40 пар. Зато органы чувств у метасприггины те же самые: вынесенные вперед камерные глаза, расположенные между ними маленькие обонятельные капсулы, а позади — капсулы для внутреннего уха. Обонятельное отверстие, скорее всего, непарное. Вдоль туловища обнаружены следы дуг позвонков. Жаберных щелей было шесть пар — тоже как у хайкоуихтиса (рис. 5).
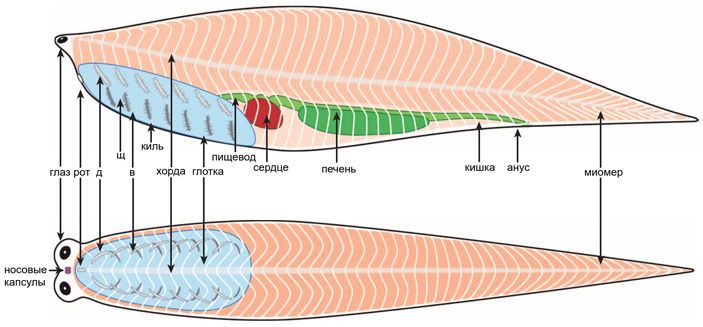
Рис. 5. Реконструкция метасприггины сбоку и с брюшной стороны. Сокращения: д — верхняя (дорсальная) часть скелета жаберной дуги, в — нижняя (вентральная) часть скелета жаберной дуги, щ — жаберная щель. Иллюстрация из статьи S. Conway Morris, J.-B. Caron, 2014. A primitive fish from the Cambrian of North America
Конвей Моррис и Карон особо подчеркивают, что границы миомеров у метасприггины образуют сильно выраженные углы. Согласно гипотезе Лакалли (с которой они согласны) это должно означать, что метасприггина, в отличие от пикайи, была хорошим и быстрым пловцом — как и подобает уважающему себя позвоночному.
Есть еще кембрийское ископаемое под названием Emmonsaspis, которое было открыто аж в XIX веке, но долго оставалось на периферии внимания палеонтологов. Конвей Моррис и Карон предположили, что эммонсаспис — синоним метасприггины; это еще нельзя считать доказанным, но в любом случае разница между ними не очень велика. Вот, собственно, и все кембрийские позвоночные, более или менее надежно известные к 2024 году.
Нуучихтис
Летом 2024 года палеонтологи Руди Лерозе-Обрил (Rudy Lerosey-Aubril) и Хавьер Ортега-Эрнандес (Javier Ortega-Hernández) опубликовали описание кембрийской окаменелости, которая много лет хранилась в отделе палеонтологии Музея естественной истории штата Юта (Natural History Museum of Utah). Это оказалось небольшое морское позвоночное. Исследователи дали ему родовое название Nuucichthys, первая часть которого произведена от самоназвания индейцев юта, а вторая от греческого слова, обозначающего рыбу. Нуучихтис очень похож на метасприггину, тоже открытую на территории Северной Америки. Но у него есть и уникальные признаки, причем весьма интересные (рис. 6).
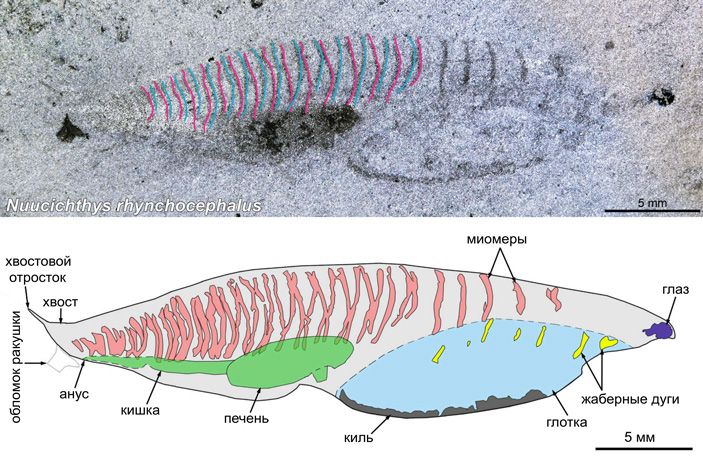
Рис. 6. Nuucichthys. Сверху — фотография отпечатка, снизу — сделанный по этому отпечатку рисунок. И на фотографии, и на рисунке видна особенность нуучихтиса, которая не обсуждается в статье, но на которую обратил внимание Александр Николаевич Кузнецов: в задней половине туловища миомеры (а вернее, миосепты) расположены как бы вдвое чаще. Скорее всего это проявление симметрии скользящего отражения, когда правые и левые сегментарные структуры сдвинуты относительно друг друга примерно на полсегмента. Именно так расположены миомеры у ланцетника: левый ряд миомеров сдвинут вперед относительно правого, и чем дальше назад, тем этот сдвиг сильнее. Ланцетник — далеко не единственный пример, симметрия скользящего отражения, она же метамерия сдвига, распространена у хордовых довольно широко (см. A. N. Kuznetsov, 2023. Glide-reflection symmetry in deuterostomes: an evolutionary perspective). На фотографии отпечатка нуучихтиса А. Н. Кузнецов пометил цветом контуры миосепт, которые, по его предположению, располагались со сдвигом: в каждом сегменте «красная» миосепта (вероятно, левая) выдвинута вперед по отношению к «синей» миосепте (вероятно, правой). Или наоборот — по двумерному отпечатку это определить трудно. В любом случае похоже, что по этому признаку нуучихтис напоминал ланцетника. Иллюстрации из обсуждаемой статьи в Royal Society Open Science, с изменениями
Нуучихтис — существо длиной 3,2 сантиметра, характерной обтекаемой формы, которую авторы статьи называют торпедообразной. У переднего конца головы хорошо заметны глаза, причем у нуучихтиса они относительно больше, чем у метасприггины. Вдоль тела расположены примерно 25 миомеров — несколько меньше, чем у метасприггины. Интересно, что границы миомеров у нуучихтиса почти вертикальны: это видно на изображениях, и авторы статьи сами характеризуют миомеры как «прямые или слегка изогнутые назад» (рис. 7). Если верны приведенные выше рассуждения Лакалли, Конвея Морриса и Карона, это должно означать, что нуучихтис был достаточно медленным пловцом (в статье этого вывода нет, но исходя из сказанного он прямо напрашивается). По данным Лакалли, у ланцетника угол наклона миосепт увеличивается с возрастом: если у личинки он составляет от 40 до 55°, то у взрослого ланцетника — от 64 до 68° (T. Lacalli, 2012. The Middle Cambrian fossil Pikaia and the evolution of chordate swimming). Поскольку нуучихтис почти вдвое меньше метасприггины, то не стоило бы исключать возможность, что и у него слабый наклон миосепт служит признаком молодого животного. Авторы обсуждаемой статьи характеризуют вероятный образ жизни нуучихтиса как «жизнь в толще воды с ограниченными способностями к плаванию или, другими словами, планктонектонный стиль жизни». С этим можно только согласиться.
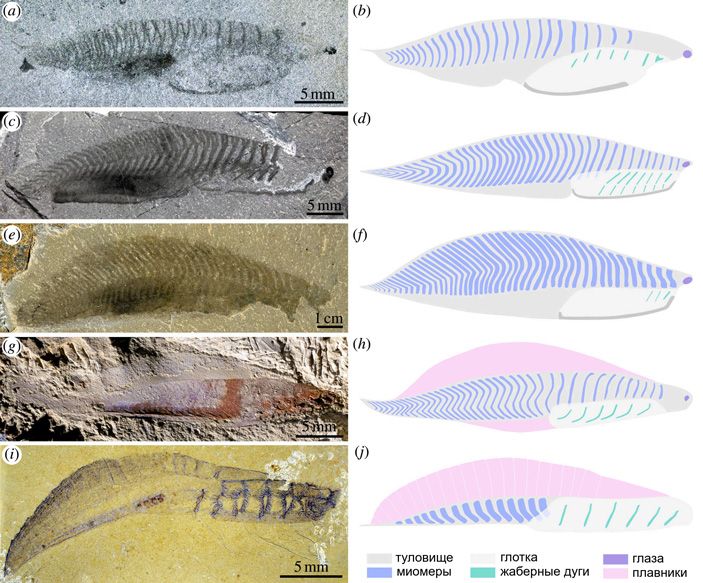
Рис. 7. Сравнение формы тела и системы миомеров: a, b — Nuucichthys, c, d — Metaspriggina, e, f — Emmonsaspis, g, h — Myllocunmingia, i, j — Yunnanozoon. Хорошо видно, что у нуучихтиса ряд миомеров не доходит до переднего конца тела. Кроме того, у метасприггины, эммонсасписа и миллокунмингии хорошо заметно, что углы наклона миосепт увеличиваются в задней половине тела, которая, собственно, в основном и создает усилия при плавании. Иллюстрация из обсуждаемой статьи в Royal Society Open Science
В нижней передней части тела нуучихтиса, как и положено, находится объемистая глотка, в которой видны остатки скелетных жаберных дуг. Последних удалось насчитать семь; значит, находившихся между ними жаберных щелей было шесть. Особый интерес представляет самая передняя жаберная дуга (рис. 8). Она расположена не только впереди всех других жаберных дуг, но, судя по всему, даже впереди от отверстия рта. Ее форма резко отличается от формы любой другой жаберной дуги; эта жаберная дуга — единственная, у которой отсутствуют вентральные (нижние) отделы. Это описание в точности подходит к предчелюстной дуге позвоночных животных, существование которой давно «вычислено» сравнительной анатомией. Очевидно, это она и есть. Следующая жаберная дуга, таким образом, обязана быть челюстной дугой. Мы видим, что форма челюстной дуги у нуучихтиса ничем принципиально не отличается от формы обычных жаберных дуг, следующих за ней. Нуучихтис — воистину бесчелюстное существо. Причем в этом отношении он гораздо архаичнее современных миног и миксин, у которых челюстная дуга хоть и не превращена в челюсти, но все равно до неузнаваемости видоизменена.
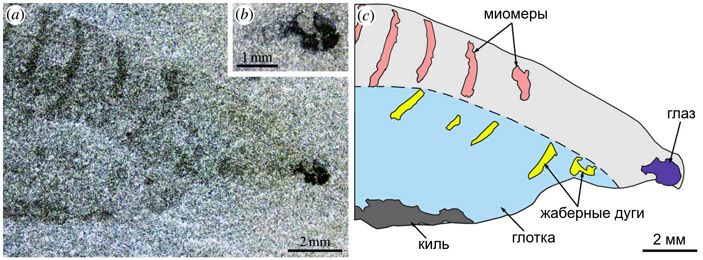
Рис. 8. Детали строения нуучихтиса: a — передняя часть тела, b — глаз, c — рисунок передней части тела. Пояснения в тексте. Иллюстрация из обсуждаемой статьи в Royal Society Open Science
Важнейшее отличие нуучихтиса от метасприггины состоит вот в чем. У метасприггины миомеры, постепенно уменьшаясь, доходят до самого переднего конца тела, как это и должно было бы быть у древнейших позвоночных. Лента миомеров у метасприггины почти достигает глаз, никакого свободного пространства впереди от миомеров там нет. А вот у нуучихтиса это не так. Миомеры у него заканчиваются существенно не доходя до переднего конца тела, и между передними миомерами и глазами есть свободное пространство. Самый передний миомер нуучихтиса находится над той жаберной дугой, которая расположена позади челюстной дуги (она называется гиоидной или подъязычной дугой). Над челюстной и предчелюстной дугами у нуучихтиса никаких миомеров не видно. Куда они делись? Вообще говоря, судьба челюстного и предчелюстного миомеров у позвоночных хорошо известна. Из них образовались внешние мышцы глаза (у большинства современных позвоночных — пять из шести мышц, поворачивающих глазное яблоко), а у некоторых рыб еще и подчерепная мышца, связывающая передний и задний блоки мозговой коробки. Можно ли уверенно утверждать, что у нуучихтиса начался распад передних миомеров, ведущий к формированию этих мышц? Пока нельзя: данные слишком неполны. Но повод для раздумий это дает серьезный. Авторы статьи справедливо замечают, что нуучихтис документирует очередной этап эволюционного процесса цефализации, формирования головы как отдела тела, отличающегося от туловища. Получается, что у нуучихтиса голова формально длиннее, чем у метасприггины.
Еще одна интересная черта нуучихтиса — отсутствие плавников, даже хвостового и спинного. Если бы эти плавники у него были, то при данных условиях захоронения их отпечатки сохранились бы наверняка (это, как и многое другое, проверялось тафономическими экспериментами — захоронениями миног, миксин и других современных животных). Здесь, таким образом, мы сталкиваемся с проблемой происхождения непарных плавников. У хайкоуихтиса непарные плавники есть, а вот у метасприггины, к которой нуучихтис ближе, они не обнаружены. Палеонтологи думали было, что это артефакт сохранности, но нуучихтис повышает вероятность того, что ни у него, ни у метасприггины плавников действительно не было. Авторы обсуждаемой статьи, во всяком случае, считают, что нуучихтис, метасприггина и за компанию с ними эммонсаспис были истинно бесплавниковыми формами. Биомеханические расчеты, основанные на версии, что метасприггина не имела хвостового плавника, показывают, что ее форма тела плохо подходит для долгого равномерного плаванья, но хорошо — для быстрых резких рывков: стремительно уплыть от хищника и затаиться (D. Rival et al., 2021. Fish without tail fins—exploring the function of tail morphology of the first vertebrates). Получается, что в кембрии сосуществовали позвоночные, использовавшие разные способы движения — плавниковые формы (хайкоуихтис и его родственники) и бесплавниковые (метасприггина и ее родственники). Бесплавниковые формы проиграли это состязание: начиная с ордовика хотя бы непарные плавники есть у всех известных позвоночных.
Подводя итог, можно сказать, что открытие нуучихтиса вряд ли станет переворотом в зоологии, но наверняка добавит несколько важных фрагментов в «пазл» происхождения позвоночных, который десятилетие за десятилетием тщательно собирают палеонтологи разных стран. Так познание эволюции в основном и движется.
Источник: R. Lerosey-Aubril, J. Ortega-Hernández. 2024. A long-headed Cambrian soft-bodied vertebrate from the American Great Basin region // Royal Society Open Science. 2023. DOI: 10.1098/rsos.240350.
Сергей Ястребов, Элементы.